
Posted: 10 Apr 2013 12:00 AM PDT
[From the site Global Warming and Teraforming. While I do not necessarily agree with this article and I count this as a guest blog, I think it is worthwhile to pass along the conclusion made at this blog article-DD]
[From the site Global Warming and Teraforming. While I do not necessarily agree with this article and I count this as a guest blog, I think it is worthwhile to pass along the conclusion made at this blog article-DD]
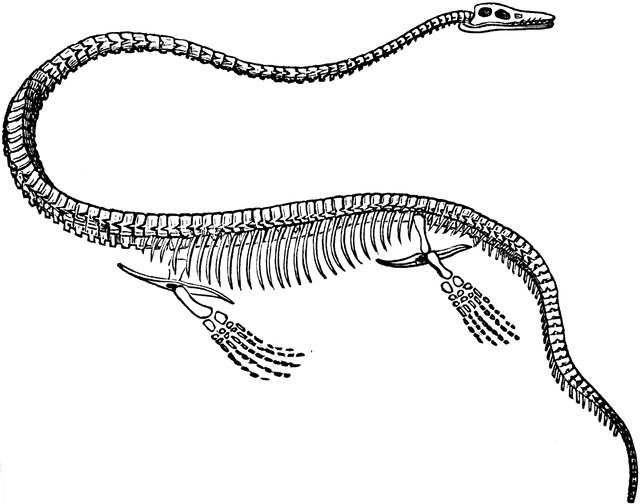
This skeleton and a recent report posted in which the creature was outright eyeballed directly from above by trained observers pretty well clinches it. Some of the inferences here are open to controversy, but what we are dealing with can be put to rest.
All sea serpent reports and loch Nessie like reports are about this creature and no other.
I think that it still retains the capacity to crawl along on land when it needs to. It certainly will be able to flounder around in a marsh. Thus establishing an egg nest in decaying vegetation is practical. While the argument for a live birth is attractive and possible, the observed return to the deep lakes make it somewhat unlikely. I also think that the creature does wait for the eggs to hatch before returning with its brood to the sea. All in, parsing a wide range of eye witness reports, the weight of evidence is describing this particular creature.
I also expect that this creature prefers the deep and is well below any hardware we deploy. It certainly avoids the sound of any shipping. Thus while our traffic has expanded exponentially, observation has plummeted during the past century.
The take home is that this creature is globally distributed and populous. It is not aggressive outside its habitat and its food must handily fit into its mouth to be held by long teeth. This is ideal for fish.
We also note that a comb like crest has been observed and that this is likely an external gill.

Elasmosaurus
Name: Elasmosaurus (Ribbon lizard)
Phonetic: El-lazz-mo-sore-us.
Named By: Edward Drinker Cope - 1868.
Named By: Edward Drinker Cope - 1868.
Classification: Chordata, Reptilia, Sauropterygia, Plesiosauria, Elasmosauridae.
Species: E. platyurus (type).
Diet: Piscivore.
Size: 14 meters long.
Known locations: USA, particularly associated with Kansas - Pierre Shale.
Time period: Campanian of the Cretaceous.
Fossil representation: Many specimens.
http://www.prehistoric-wildlife.com/species/e/elasmosaurus.html
Today Elasmosaurus is one of the best known plesiosaurs in the world, and is the type genus for the Elasmosauridae. This group is noted for having particularly long necks, proportionately even longer than the more standard plesiosaurs. These long necks are composed of a large number of cervical vertebrae that in Elasmosaurusnumber seventy-one, more than other plesiosaurs including fellow elasmosaurids. The sheer number of vertebrae not only extend the length of the neck but also provide greater flexibility for fine control of the head, although very early reconstructions of Elasmosaurus being able to coil its neck like a snake are today considered to be very inaccurate and impossible for a living Elasmosaurus to do. Instead the neck would have been held quite straight when resting with sideward movements only occurring when necessary.
It was the long neck of Elasmosaurus that was key to its feeding method. All Elasmosaurus would have to do was swim up to a shoal of fish, possibly from below so that it could hide its body in the slightly darker depths, and use its neck to dart its head in and pluck out a mouthful of fish. Approaching prey like a shoal of fish from below would also silhouette themselves against the brighter surface water, making them easier to define as well as removing the defence method of flashing light off their shiny scales to confuse predators as they turned and swam through the water. Such a method would have been a reasonably efficient form of feeding as Elasmosaurus would just have to move its head and neck and not waste energy swimming after fast moving individuals.
Key to prey capture were the long and thin teeth that protruded from the mouth of Elasmosaurus. These intermeshed together so that when a fish was caught between the jaws it was impaled upon the teeth so that it could not wriggle free and escape. This kind of dentition is common among other elasmosaurids as well as other piscivorous(fish eating) animals such as skim feeding pterosaurs. A defining characteristic of Elasmosaurus is the fact that it has six teeth in each premaxilla. Because the teeth of Elasmosaurus were adapted for prey capture rather than biting clean through flesh, prey would need to have been swallowed whole. Gastroliths have been found in association to Elasmosaurusremains and its thought that the grinding action of these stones in its gut tenderised food for easier digestion.
Elasmosaurus probably was not a fast swimmer, although if it used stalking tactics to approach its prey from below it would not need to be. The flipper-like limbs were rigid and well suited for providing efficient locomotion through the water. Skeletal elements of the main body such as the pectoral girdles were also well adapted to provide efficient musculature support for the flippers so that that Elasmosaurus could paddle itself through the water with ease.
While Elasmosaurus was beautifully adapted for an aquatic life, the price of these adaptations is that Elasmosaurus would have almost certainly been restricted to the marine environment. Because the paddle-shaped limbs were so stiff they would at best been only just better than useless for pushing its body along. Even when if Elasmosaurus was ever active in the shallows, it would still need to have kept its body totally immersed so that the water could support its body weight.
More ‘classic’ depictions of Elasmosaurus shooting its neck out of the water are also unlikely as the longer the neck got, the more body tissue such as bone and muscle was used to make up the neck meaning more weight was added to it. More weight means that stronger muscles would have been needed to support the neck when out of the water, and study of the cervical vertebrae where the muscles attach indicates that Elasmosaurus just did not have the muscles to move its neck when it did not have water to support the weight. Instead the muscles were only strong enough to provide basic neck movements, which would be all that was required of them when the neck was immersed in water.
While no hard evidence currently exists for Elasmosaurus either laying eggs on land or giving birth to live young, most palaeontologists believe that Elasmosaurus, and similar marine reptiles would have given birth to live young. The precedent is known to exist for other marine reptiles like the ichthyosaurs and nothosaurs as far back as the Triassic and given the problems that Elasmosaurus would have had just moving its body if it ever found itself out of the water make it extremely unlikely it could have laid eggs on land. Today marine reptiles such as sea snakes can also be observed giving birth to live young which indicates that as a group the reptiles are capable of adapting to new methods to thrive in different environments.
Elasmosaurus discovery, reconstruction and classification
The palaeontological history of Elasmosaurus is a turbulent one. First discovered in 1868 by Dr. Theophilus Turner, the fossils were sent to Edward Drinker Cope for study and identification. Cope had never assembled a plesiosaur before but he had the idea the large number of vertebrae mostly belonged in the tail which resulted in a long tail and short neck. To Cope this made sense as the proportions were similar to those of many modern lizards that also have short necks and incredibly long tails.
The next part of this story is a little hazy for exact details, but the popular retelling of it has Othniel Charles Marsh realising that the head was on the wrong end and ridiculed Cope for placing it in the wrong place. However it seems that it was actually Joseph Leidy who pointed this error out in a meeting in 1870, although given the animosity already between Marsh and Cope at the time, its quite possible that Marsh continued to use this case as an example to discredit Cope’s work, hence the confusion. It is this criticism that poured fuel onto the already burning fire that would become the ‘bone wars’, a fierce rivalry between Cope and Marsh that saw them trying to out compete one another in terms of discovering new species.
Since its naming back in 1868 many new specimens of Elasmosaurus have been recovered, and many of these were assigned their own species within the genus. Many of these species continue to be mentioned despite the fact that in 1999 the palaeontologist Ken Carpenter conducted a study of the known Elasmosaurus fossils at this point and found that some of these did not belong as individual species, and some of them even represented different genera. Today most palaeontologists only recognise the original type species, E. platyurus, as being valid.
No comments:
Post a Comment
This blog does NOT allow anonymous comments. All comments are moderated to filter out abusive and vulgar language and any posts indulging in abusive and insulting language shall be deleted without any further discussion.