

Recent model of the likely surfacing configuration taken on by Cryptoclidus compared to the Devon Coast, Nushagak Bay, and Pembroke Dock 'sea serpent' photographs

FRONTIERS OF ZOOLOGY
Dale A. Drinnon has been a researcher in the field of Cryptozoology for the past 30+ years and has corresponded with Bernard Heuvelmans and Ivan T. Sanderson. He has a degree in Anthropology from Indiana University and is a freelance artist and writer. Motto: "I would rather be right and entirely alone than wrong in the company with all the rest of the world"--Ambroise Pare', "the father of modern surgery", in his refutation of fake unicorn horns.



The Mary River turtle (Elusor macrurus) with a green mohawk of algae.
It's suggested that the algae helps provide camouflage.
Jay Cooney just sent me a copy of the photo and made the suggestion
Apparently the turtles do this as camouflage. A longneck doing this could definitely explain the seaweed-like "hair"[in the "mane"]!
And actually that is a quite good explanation. the seaweed (algae) would also be growing in seasonally, in both saltwater and fresh (with different species) and also could look either greenish or reddish brown, and so it fits all of the broad criteria. Furthermore, the animals could be tearing it off of each other by mouth without doing each other any harm, and the photos do show other patches of growth on the face in the area where "ear fins" and "whiskers" [even "green whiskers"] are rarely and irregularly reported
So I am going to pencil that in a strong possibility and in fact I now rank it as a strong possibility, a higher possibility than Bernard Heuvelmans' and Ivan Sanderson's suggestion that vascularized fibers make up the "Mane." I do not have it as the default yet but it might well be the explanation that stands the Occam's Razor test the best.
Best Wishes, Dale D.
|









 |
| water_horse_by_giniwolf-d4sazyp |

















 |
| Umfuli SS, after Captain Cringle |






 During the following winter, Freeman and other investigators found additional tracks on several occasions. They made more plaster casts indicating the existence of at least two more individuals. In all, 11 casts of four distinguishable types have been made. Five of these casts were made by Forest Service workers in June of 1982, and the originals are still in my possession, pending their ultimate disposition. One made by Art Snow of the search and rescue team, and five more made by Paul Freeman himself, have been copied with latex or Silastic molds and returned to their owners.
During the following winter, Freeman and other investigators found additional tracks on several occasions. They made more plaster casts indicating the existence of at least two more individuals. In all, 11 casts of four distinguishable types have been made. Five of these casts were made by Forest Service workers in June of 1982, and the originals are still in my possession, pending their ultimate disposition. One made by Art Snow of the search and rescue team, and five more made by Paul Freeman himself, have been copied with latex or Silastic molds and returned to their owners. sizes of around 0.01 mm diameter. When this soil is damp and cool, and pressed into by a warm body, a detailed imprint commonly remains. Most, if not all, of these tracks were cast within a day of the time of impression, so they had not dried out, nor had they received any intrusive material. The well mixed casting plaster that was poured into these impressions was able to record any degree of detail that was held by the soil. Variations down to less than 0.1 mm are faithfully preserved. This is fine enough to show individual dermal ridges and their sweat pores.
sizes of around 0.01 mm diameter. When this soil is damp and cool, and pressed into by a warm body, a detailed imprint commonly remains. Most, if not all, of these tracks were cast within a day of the time of impression, so they had not dried out, nor had they received any intrusive material. The well mixed casting plaster that was poured into these impressions was able to record any degree of detail that was held by the soil. Variations down to less than 0.1 mm are faithfully preserved. This is fine enough to show individual dermal ridges and their sweat pores. than the investigator's boots, in spite of the fact that they covered over twice the area of these boots. At the location of Freeman's alleged sighting, the Forest Service conducted a test at the urging of an individual visiting from Seattle. A piece of metal the size and shape of one of the footprints was cut, and this was placed on the dirt road near a track. An auto jack was then put on it, and this was used to raise the rear end of a pickup truck. They found that this metal plate was impressed into the ground less that the actual footprints. From this fact, and given the weight of the half supported truck, the Seattle individual concluded the "thing" weighed 4,000 pounds (1914 kg). None of the foresters agreed with this weight (widely quoted in the press), allowing only one-fourth of it at most.
than the investigator's boots, in spite of the fact that they covered over twice the area of these boots. At the location of Freeman's alleged sighting, the Forest Service conducted a test at the urging of an individual visiting from Seattle. A piece of metal the size and shape of one of the footprints was cut, and this was placed on the dirt road near a track. An auto jack was then put on it, and this was used to raise the rear end of a pickup truck. They found that this metal plate was impressed into the ground less that the actual footprints. From this fact, and given the weight of the half supported truck, the Seattle individual concluded the "thing" weighed 4,000 pounds (1914 kg). None of the foresters agreed with this weight (widely quoted in the press), allowing only one-fourth of it at most. This excessive weight figure is easily dismissed because it is based on a gradual pressing of weight into the ground, in contrast to the impact of striding. A simple experiment illustrates this: One can walk with long and firm strides on fine moist soil, leaving clear footprints, then stop and walk slowly back to examine the tracks that were just made. One may then backoff and compare the "striding" tracks with the "walking" tracks, and see that the latter are far less impressed. I have found that, in most cases, even hard stamping in-place will not match the impressions just previously made in striding. Full striding impacts the body weight, at some speed, entirely on the heel area initially, and with a slight forward motion as well. In leaving the ground the body weight is last impressed on the toe area, with the force of stepping off, and also with a slight backward motion.
This excessive weight figure is easily dismissed because it is based on a gradual pressing of weight into the ground, in contrast to the impact of striding. A simple experiment illustrates this: One can walk with long and firm strides on fine moist soil, leaving clear footprints, then stop and walk slowly back to examine the tracks that were just made. One may then backoff and compare the "striding" tracks with the "walking" tracks, and see that the latter are far less impressed. I have found that, in most cases, even hard stamping in-place will not match the impressions just previously made in striding. Full striding impacts the body weight, at some speed, entirely on the heel area initially, and with a slight forward motion as well. In leaving the ground the body weight is last impressed on the toe area, with the force of stepping off, and also with a slight backward motion.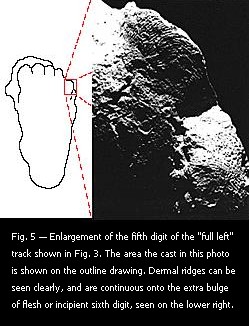 A peculiarity of the "full left" track (Fig. 5) is the extra pad of flesh to the lateral side of the base of digit V. Dermal ridges pass directly from digit V onto this pad, so it is not a double strike of the fifth digit. Since this pad does not appear on either of the "right" tracks, its exact nature is not clear. It may result from connective tissues in the sole pad that were under a normal tension. Possibly it is an incipient sixth digit.
A peculiarity of the "full left" track (Fig. 5) is the extra pad of flesh to the lateral side of the base of digit V. Dermal ridges pass directly from digit V onto this pad, so it is not a double strike of the fifth digit. Since this pad does not appear on either of the "right" tracks, its exact nature is not clear. It may result from connective tissues in the sole pad that were under a normal tension. Possibly it is an incipient sixth digit. that was also very flexible. Mobility of bone connections cannot account for most of this indentation. The deepest part is close enough to the back of the foot that only the calcaneous, or heel bone, is involved. It is obvious that the sole pad must have had a thickness somewhat greater than the 2 cm depth of the stone's indentation. This applies to the osteologically more rigid rear part of the foot, and may be much the case in the forefoot, where there is more movability between the bones.
that was also very flexible. Mobility of bone connections cannot account for most of this indentation. The deepest part is close enough to the back of the foot that only the calcaneous, or heel bone, is involved. It is obvious that the sole pad must have had a thickness somewhat greater than the 2 cm depth of the stone's indentation. This applies to the osteologically more rigid rear part of the foot, and may be much the case in the forefoot, where there is more movability between the bones. body weight will be distributed over about 200² cm of one foot (at times), or about 0.325 kg per cm². Our reconstructed Sasquatch, 2.4 - 2.5 m tall, has a foot 37.5 cm long. Its 400 kg body weight is distributed over 486² cm (actual measure) of one foot, or about 0.822 kg per cm².
body weight will be distributed over about 200² cm of one foot (at times), or about 0.325 kg per cm². Our reconstructed Sasquatch, 2.4 - 2.5 m tall, has a foot 37.5 cm long. Its 400 kg body weight is distributed over 486² cm (actual measure) of one foot, or about 0.822 kg per cm².  Such a thick sole pad would presumably be made largely of fat, but would have to be strongly interlaced with, and partitioned by, connective tissue running in many directions. This design would allow for the retention of a typical higher-primate friction-skin surface with a minimum of internal restructuring. This conclusion, which was based only on Sasquatch footprint morphology and estimated body sizes, is supported by data on the gorilla foot. Raven (1950: 217) describes the gorilla sole as follows: "The subcutaneous connective tissue ... is very thick and fatty and similarly has a framework of heavy collagenous septa." And (1950: 71): "The fatty pad of the sole is everywhere covered by dense connective tissue. The fat is thin along the middle of the foot but is very thick, 2.5 cm under the heel." Given that the Sasquatch may weigh twice as much as the gorilla, and is entirely bipedal, it is reasonable to expect the same sole structure, but to an exaggerated degree.
Such a thick sole pad would presumably be made largely of fat, but would have to be strongly interlaced with, and partitioned by, connective tissue running in many directions. This design would allow for the retention of a typical higher-primate friction-skin surface with a minimum of internal restructuring. This conclusion, which was based only on Sasquatch footprint morphology and estimated body sizes, is supported by data on the gorilla foot. Raven (1950: 217) describes the gorilla sole as follows: "The subcutaneous connective tissue ... is very thick and fatty and similarly has a framework of heavy collagenous septa." And (1950: 71): "The fatty pad of the sole is everywhere covered by dense connective tissue. The fat is thin along the middle of the foot but is very thick, 2.5 cm under the heel." Given that the Sasquatch may weigh twice as much as the gorilla, and is entirely bipedal, it is reasonable to expect the same sole structure, but to an exaggerated degree. The near constancy of ridge spacing in primates rules out one method of faking. Latex molds of real skin, soaked in kerosene, will expand greatly; there may also be other methods of expanding molds. This procedure could produce gigantic skin patterns (in some respects), but the ridge spacing would also be expanded, and thus easily recognizable as abnormal.
The near constancy of ridge spacing in primates rules out one method of faking. Latex molds of real skin, soaked in kerosene, will expand greatly; there may also be other methods of expanding molds. This procedure could produce gigantic skin patterns (in some respects), but the ridge spacing would also be expanded, and thus easily recognizable as abnormal.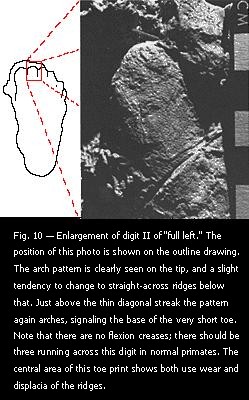 growth progresses, these cones line up in fixed patterns and merge together into unbroken ridges. Sometimes, this development partially fails, and many of the ridges consist only of a jumble of very short segments. This condition is known as displacia (or dysplasia), and tends to affect mostly those central areas of greatest weight support where the ridges are also most worn down. While the two conditions topographically coincide, they should not be confused, because they have separate origins and morphology. In affected areas, the directions of dermal ridge sets can still be made out by observing the general trend of orientation of the short segments.
growth progresses, these cones line up in fixed patterns and merge together into unbroken ridges. Sometimes, this development partially fails, and many of the ridges consist only of a jumble of very short segments. This condition is known as displacia (or dysplasia), and tends to affect mostly those central areas of greatest weight support where the ridges are also most worn down. While the two conditions topographically coincide, they should not be confused, because they have separate origins and morphology. In affected areas, the directions of dermal ridge sets can still be made out by observing the general trend of orientation of the short segments. flexion creases (see especially Fig. 10). Ideally, there should be three such creases, each corresponding to one of the three phalangeal joints, but in this case none are visible. I do not know how to interpret this absence, except to make note of it. It may not be a normal Sasquatch trait, because the toes on this particular individual are relatively even shorter than on other presumed Sasquatch footprints. The main significance, for now, may relate to the possibility of these tracks being faked with lifts and casts from real primate skin. How does one locate a group of people with arched thumbs of various sizes, and uniformly short, with none of them showing any flexion creases? While there are many pattern breaks with large blank areas on the soles, some of the toe prints are continuous from tips to metatarsal pads. No patching is indicated. I could find only one case in the literature of a human finger without flexion creases. Haylock (1983) illustrates one of a brachydactylous individual, where a very short digit II lacks creases and has a continuous arch pattern from the tip to the metacarpal end.
flexion creases (see especially Fig. 10). Ideally, there should be three such creases, each corresponding to one of the three phalangeal joints, but in this case none are visible. I do not know how to interpret this absence, except to make note of it. It may not be a normal Sasquatch trait, because the toes on this particular individual are relatively even shorter than on other presumed Sasquatch footprints. The main significance, for now, may relate to the possibility of these tracks being faked with lifts and casts from real primate skin. How does one locate a group of people with arched thumbs of various sizes, and uniformly short, with none of them showing any flexion creases? While there are many pattern breaks with large blank areas on the soles, some of the toe prints are continuous from tips to metatarsal pads. No patching is indicated. I could find only one case in the literature of a human finger without flexion creases. Haylock (1983) illustrates one of a brachydactylous individual, where a very short digit II lacks creases and has a continuous arch pattern from the tip to the metacarpal end. The possibility that air bubbles might have mimicked sweat pores was suggested by physical anthropologist Tim White, at the University of California, Berkeley, who otherwise thought the casts appeared to represent legitimate footprints. To settle this point, I made impressions of false ridges (with a fine comb) in similar soil, and cast them in plaster. I compared the results with the actual casts, and found that there are, in fact, occasional air bubbles from casting. These bubbles, however, are sharp-edged, and are not as small as the apparent sweat pores. They are rather few, and not regularly spaced or lined up. In some cases, they also bulge out the ridges around them, but only slightly, and with a much thinner wall between the hole and the ridge edge than with the presumed pores (Fig. 13).
The possibility that air bubbles might have mimicked sweat pores was suggested by physical anthropologist Tim White, at the University of California, Berkeley, who otherwise thought the casts appeared to represent legitimate footprints. To settle this point, I made impressions of false ridges (with a fine comb) in similar soil, and cast them in plaster. I compared the results with the actual casts, and found that there are, in fact, occasional air bubbles from casting. These bubbles, however, are sharp-edged, and are not as small as the apparent sweat pores. They are rather few, and not regularly spaced or lined up. In some cases, they also bulge out the ridges around them, but only slightly, and with a much thinner wall between the hole and the ridge edge than with the presumed pores (Fig. 13). depth of imprinting are also beyond human capability. Human feet do not have the flexibility of sole to conform to the large stone that was stepped on in one case. The toe design is also beyond the range of human feet in that they are relatively too short, toes II through V are too wide, and they line up too nearly straight across the front of the foot.
depth of imprinting are also beyond human capability. Human feet do not have the flexibility of sole to conform to the large stone that was stepped on in one case. The toe design is also beyond the range of human feet in that they are relatively too short, toes II through V are too wide, and they line up too nearly straight across the front of the foot.I see dermal ridges of the arch type distally directed. I see sweat pores. If it's a fake, it's a brilliant fake, on the level of counterfitting, and by someone well versed in dermatoglyphics.Anthropologists Mikhail Urisson and Vladimir Volkov-Dubrovin (Deputy Director of the Institute) agreed with the above opinion.
Regarding photographs of skin imprints sent over by G. Krantz. I fully agree with his opinion on these footprints, as well as the opinion of Benny Kling. The structure of the dermal ridges is very much like that of man. The sweat glands have large openings because the ridges are much bigger than in man. It was great luck that the footprints were left in the soil that revealed fine details of the imprints. As for the patterns of ridges, some irregularity in ridge lines in separate places in the photos may be connected with the peculiarity of the material in which the imprints were made (unevenness of soil, various inclusions, such as small pebbles, pine needles, etc.). Another possibility is scars and skin injuries.Douglas M. Monsoor, Supervisor, Criminalistics Unit, Department of Public Safety, Lakewood, Colorado. Certified Latent Print Examiner, and fellow of the Fingerprint Society of the United Kingdom. Was sent detailed photographs in late 1982, examined original casts in December, 1982, and again in June, 1983, for two hours on each occasion.
Incidentally, even in ideally made human imprints there can be such irregularities. There is even a whole branch of dermatoglyphics studying genetic irregularities in ridge lines, i.e. medical and genetic dermatoglyphics.
In the imprints shown by the available photographs, I cannot detect anything unusual, except digit I, left foot, which shows, apparently, a pattern of the arch type (in man the whorl type is more frequently found).
I see the presence of ridge structure in these track casts which, in my examination, appears consistent with that type of ridge structure you would find in a human. Under magnification, they evidence all the minute characteristics similar to human dermal ridges. The sizes, distributions, and orientations of the ridge patterns are consistent with those found on a human foot. Of the ridge structure visible in the impressions, I believe it was produced concurrent with the creation of the overall impressions, and not added later.Robert D. Olsen, Sr., Criminalist, Kansas Bureau of Investigation, Topeka, Kansas. Certified Latent Print Examiner, Fellow of the American Academy of Forensic Sciences, Fellow of the Fingerprint Society of the United Kingdom, Member of International Association for Identification, etc. Was sent detailed photographs in late 1982, and Silastic lifts in early 1983. Examined original casts and two-color lifts in June, 1983, for several hours.
If hoaxing were involved, I can conceive of no way in which it could have been done. They appear to be casts of original impressions of a primate foot — of a creature different from any of which I am aware.
Based on everything I see, there is nothing in these tracks that is inconsistent with the impressions of an actual living primate foot. Ridges and pores are consistent with real primate skin. I'm convinced that this represents real friction skin and shows no inconsistencies in structure or orientation.Edward Palma, Fingerprint examiner for the Laramie County Sheriff's Department, Cheyenne, Wyoming. Had latex lifts from the footprints in late 1982 — inked these and traced print pattern extensively. Had cast copies to examine at leisure. Saw original casts for several hours in early 1983 — made Duplicast impressions of critical parts, and took photographs for further study.
If they are faked, the individual would have to know an extraordinary amount about fingerprinting. I could not have done it. A faker would have to be an accomplished artist as well as an expert on dermatoglyphics. He would also need a knowledge of gross anatomy of feet. The amount of time needed to do all this work is beyond the realm of believability.
My professional opinion of the three casts is that they represent footprints of a living higher primate of an unknown species. The over-all configuration of the foot is roughly human, but it is too wide — a human foot would not be over five and one half inches [wide] for this length, and thus these impressions could not be human. The actual width is represented and supported by ridge pattern.Benny Kling, Instructor, Law Enforcement Academy, Douglas, Wyoming. Had latex lifts from the footprints in late 1982, and cast copies shortly thereafter. Saw original casts for several hours on two occasions in early 1983.
My study of the tracks concentrated more on the sole than on the more conspicuous details of the toes. I traced the ridge pattern over the entire breadth of the forefoot, finding triradius landmarks appropriate in their respective positions with intervening ridges flowing in proper directions. It could not have been patched together from smaller parts that were copied from skin of a known primate.
The detailed morphology of the ridge and furrow structures and patterns are especially convincing to me. In all details, they conform perfectly in design and size to real friction skin. The sweat pores are clear and are lined-up and spaced just as expectable, and can be distinguished from occasional air bubbles in the casts.
I began this investigation with the goal of showing how these prints were, or might have been, faked. All evidence now tells me that any faking would be impossible.
These track casts show all the characteristics of real friction skin derived from a higher primate footprint. The ridge details, in all respects, duplicate that found in human feet. Parts of the pattern on right and left feet are near-mirror images; some displacia is indicated in the areas where it could be expected; smoothing by wear shows on the weight-bearing areas. In addition, the footprints indicate that an unusual proportion on the body weight fell on the front of the foot, and the arches are evidently flat.
This kind of print could not have been made by a human foot, nor that of any known animal. It could not have been manufactured by any hoaxer; the design is too dermatoglyphically correct, and the engraving job would be beyond the capabilities of the best forger. Descriptions of the supposed Bigfoot, or Sasquatch, are consistent with the traits found in these footprints.
{kind=link}