PERSPECTIVES ON THE “LIVING PLESIOSAUR” CONTROVERSY Edited by and featuring commentary by Scott Mardis
“
Loch Ness Monsters (Nessies) are—if they exist—animals of a species either not yet known to science or known but thought to have been long extinct.
Much controversy has concerned eyewitness testimonies and photographs whose relevance and validity are uncertain. However, there also exists a body of objective evidence that critics have been unable to gainsay: the Dinsdale film; numerous sonar echoes obtained over many years by different investigators; and underwater photography in 1972 coincident with sonar detection of large targets.
If the descriptions of Nessies provided by photos and eyewitnesses could be interpreted as some species of animal known from anywhere else in the world, there would be no great fuss about it. If sharks, say, or dolphins, or some small whales had adapted to fresh water, that would be quite interesting to biologists but no reason for world-wide media or public interest. The trouble is, Nessies look like nothing now known to be aliveanywhere. Perhaps even worse, they look rather dinosaur-like. The real animals that they resemble most closely are plesiosaurs, marine creatures that once thrived in the oceans all over the globe but that are believed to have been extinct for tens of millions of years .”- Henry H. Bauer, The Case for the Loch Ness “Monster”: The Scientific Evidence, Journal of Scientific Exploration 16(2) (2002), pp. 225–246
Top image: 1975 Academy of Applied Science Loch Ness photograph, Bottom image: Skeleton of Cryptocleidus oxoniensis plesiosaur
“I think they got frightened. Those who make their living from this, the zoologists, are not ready to believe, on the basis of one picture, that something that should have been dead 65 million years ago is still existing in some form at Loch Ness, Scotland.” - Dr. Robert Rines, NOVA: The Beast of Loch Ness, PBS Television, 1999
"The publication of Scott and Rines and the photographs in the national press indicate that there may be a plesiosaur-like reptile inhabiting Loch Ness . It is exceedingly difficult to envisage how a former tropical marine reptile could endure the cold waters and harsh environment provided by a small lake in Scotland . Since Loch Ness did not exist until some 12,000 years ago, one is faced with the problem of the survival of Nessiteras for a period of 64 million years in a world where it’s former ecological niche has been occupied by modern cetaceans and pinnipeds” – L.B. Halstead, P.D. Goriup and J.A. Middleton, Departments of Geology and Zoology, University of Reading, U.K., letter to the journal Nature 259 (1976), pp. 75-76.
“What the body temperature and thermoregulation processes of extinct vertebrates were are central questions for understanding their ecology and evolution. The thermophysiologic status of the great marine reptiles is still unknown, even though some studies have suggestedthat thermoregulation may have contributed to their exceptional evolutionary success as apexpredators of Mesozoic aquatic ecosystems. We tested the thermal status of ichthyosaurs, plesiosaurs, and mosasaurs by comparing the oxygen isotope compositions of their tooth phosphate to those of coexisting fish. Data distribution reveals that these large marine reptiles were able to maintain a constant and high body temperature in oceanic environments ranging from tropical to cold temperate. Their estimated body temperatures, in the range from 35°
T 2°C to 39°
T 2°C, suggest high metabolic rates required for predation and fast swimming over large distances offshore.
Paladino et al. proposed that some marine reptiles such as leatherback turtles display endothermy instead of inertial homeothermy, thus helping them to feed in cold waters. However, Lutcavage et al. showed that the studied gravid female specimens raised their metabolic rates because of egg laying, thus biasing the evaluation of their true metabolic status.
Adaptation to cold marine waters was also revealed by the fossil reptile assemblage discovered in the Aptian southern high-latitude deposits of the White Cliffs in southeast Australia. The specimens were attributed to at least three families of plesiosaurs and at least one of ichthyosaurs . Paleoclimatic proxies indicate cold to near-freezing conditions at the seasonal scale, a climate mode that is not tolerated by modern ectothermic reptiles such as turtles or crocodiles. This observation suggests that some Mesozoic marine reptile taxa were able to cope with low temperature marine environments .”-Aurélien Bernard, et al. Regulation of Body Temperature by Some Mesozoic Marine Reptiles, Science 328(2010);pp.1379-1382
(from
Ryosuke Motani, Warm-Blooded “Sea Dragons”?, Science 328, 2010, pp.1361-1362)
(Left) Mesozoic Geologic Table. (Right) South Pole (Early Cretaceous Period).
“The subsurface opal-bearing deposits of the Bulldog Shale at Coober Pedy and Andamooka in South Australia, and the Doncaster Member of the Wallumbilla Formation at White Cliffs in New South Wales, represent localities of some significance, producing ichthyosaurs and a high diversity of plesiosaur taxa. The sediments comprise predominantly finely laminated shaly mudstones and claystones representing deposition under transgressive shallow marine conditions, and in an Early Cretaceous high latitude zone (60(–70(S;). Because of severe weathering and often-poor locality data (a result of specimens being discovered serendipitously and extracted during opal mining), assignment of most marine reptile fossils to exact stratigraphic horizons is again problematic. However, a predominantly Aptian age can be suggested on the basis of macro invertebrate assemblages , and the presence of potentially ice-rafted quartzite/porphyritic boulders and glendonite (pseudomorphs of the calcium carbonate hexahydratemineral ikaite) nodules
.
These conspicuous sedimentary structures characterise the early depositional stages of the Bulldog Shale and Wallumbilla Formation in the southern Eromanga Basin and have been correlated with a period of very cold to near-freezing climatic conditionsduring the Late Neocomian–Early Albian. A similar cool temperate to very cold environmental setting has been suggested for the Lower-mid Albian (C. paradoxa Zone) estuarine–coastal plain facies of the Griman Creek Formation at Lightning Ridge, New South Wales/Suratregion, Queensland, and the Aptian–Lower Albian (C. hughsiiSubzone–C. striatus Subzone) braided stream and overbank flood plain deposits of the Wonthaggi and Eumeralla formations in Victoria. These units have produced a handful of plesiosaur remains (mainly isolated teeth) that represent animals living near to or within the Cretaceous southern polar circle and evidently adapted to at least seasonal occupation of inland freshwater environments .
Leptocleidus is currently the most widely distributed plesiosaur genus known from Australia. Isolated remains also possibly attributable to Leptocleidus have been described from Albian freshwater sediments of the Griman Creek Formation at Lightning Ridge in New South Wales and the Surat district of Queensland. The bulk of these specimens consist of isolated teeth, although some skeletal elements, including a large propodial similar to that of Leptocleidus clemai (from the Hauterivian–Barremian Birdrong Sandstone of Western Australia),have been discovered. Fragmentary pliosauroid teeth and ribs closely resembling those of Leptocleidus sp. have also been recorded from the freshwater braided stream and overbank floodplain deposits of the Wonthaggi and Eumeralla formations in Victoria. Interestingly , these deposits were laid down in a very cold, high-latitude environment and provide evidence of plesiosaurs living within inland streams and rivers at the Cretaceous South Pole.”- Benjamin P. Kear, Cretaceous marine reptiles of Australia: a review of taxonomy and distribution, Cretaceous Research 24 (2003), pp. 277–303
“ The record of plesiosaurians from freshwater deposits is sparse in comparison to those from marine sediments. Despite this, a number of discoveries have been made from around the world. The fact that these range in age from Early–Middle Jurassic to Late Cretaceous attests to the group’s long history of habitation in non-marine environments. The vast majority of non-marine plesiosaur specimens are fragmentary, and many are taxonomically uninformative. Where they are diagnostic, however, many of the freshwater specimens are referable either to ‘rhomaleosaurid-like’ taxa, or to the widespread Cretaceous pliosauroid genus Leptocleidus. Not surprisingly, therefore, the material from southeastern Australia shares similarities with this latter taxon, and lends support to the hypothesis that freshwater and near-shore marine environments may have served as refugia for plesiomorphic pliosauriform plesiosaurs well into the late Early Cretaceous. The plesiosaur fossils from southeastern Australia constitute one of a number of recognized finds from Cretaceous high-latitude deposits. However, most other occurrences are marine in origin, including examples from central Australia, New Zealand, the Chatham Islands, Patagonia, Antarctica and the Canadian Northwest Territories. Amongst the currently documented specimens, those from the Early Cretaceous units of central and southeastern Australia are unusual because they occur in association with paleoclimatic indicators (e.g., cryoturbated sediments, glacial erratics, glendonites, and growth-banded wood) denoting seasonally very cold to near freezing conditions. This contrasts markedly with climatic regimes typically tolerated by modern aquatic reptiles, but suggests that some plesiosaur taxa may have been able to cope with extremely low average water temperatures.”-Benjamin P. Kear, PLESIOSAUR REMAINS FROM CRETACEOUS HIGH-LATITUDE NON-MARINE DEPOSITS IN SOUTHEASTERN AUSTRALIA, Journal of Vertebrate Paleontology 26(1) (2006), pp.196–199

Freshwater plesiosaurs from southeastern Australia (from Kear 2006)
“The taxonomic status of three previously reported plesiosaurian specimens from the non-marine Horseshoe Canyon Formation (Campanian-Maastrichtian, Upper Cretaceous) is reassessed. The holotype of Leurospondylus ultimus and a previously undescribed partial skeleton from Red Deer River Valley represent indeterminate elasmosaurids. The two plesiosaurian specimens came from different horizons for which different environments are inferred. The larger individual may be closely related to
Elasmosaurus . This study supports the hypothesized decline of polycotylid plesiosaurs in the North American during the Late Campanian and suggests small adults and juveniles of elasmosaurids existed in both marine and non-marine environments whereas large adults were limited to the latter .”- Tamaki Sato and Xiao-Chun Wu,REVIEW OF PLESIOSAURIANS (REPTILIA: SAUROPTERYGIA) FROM THE UPPER CRETACEOUS HORSESHOE CANYON FORMATION IN ALBERTA, CANADA,Paludicola 5(4) (2006), pp.150-169
Freshwater plesiosaurs, probably Elasmosaurid, from Horseshoe Canyon, Alberta, Canada(from Sato and Wu 2006)
“While the majority of plesiosaur material is associated with marine shelf deposits, some specimens have been found associated with lagoonal, estuarine or delta-margin environments and freshwater environments. Sedimentological evidence for the environment of deposition of the Axel Heiberg locality indicates that it was deposited in a lagoon or bay, but the evidence is equivocal as to the salinity of the water during deposition. While the occurrence of the plesiosaur suggests marine or brackish-water conditions, the associated fauna suggests a non-marine environment. Taxa found in association with plesiosaurs recovered from marine and near-shore lagoonal environments typically include a diverse assemblage of marine vertebrates. Mosasaurs (Kear, 2003) are typically found in association with plesiosaurs in Late Cretaceous vertebrate assemblages. Other marine vertebrates typically found in association with plesiosaurs include marine turtles and various fishes, such as Enchodus, Coelodus,Oseroides, and Hoplopteryx. However, such taxa are absent in the Axel Heiberg assemblage. At this locality, the vertebrates found in association with the plesiosaur remains are all taxa that are typical of non-marine vertebrate assemblages.Thus the faunal evidence suggests that these teeth represent an additional non-marine occurrence of plesiosaurs.
Plesiosaur occurrences in non-marine settings are typically dominated by small individuals. In the Late Campanian Dinosaur Park Formation of Alberta, all the plesiosaur remains recovered are from individuals of small size, and one-third of the specimens show features indicative of juvenile individuals. The small size of the plesiosaur from the Axel Heiberg assemblage is consistent with the dominance of juveniles in non-marine environments. The Cretaceous saw an explosion of plesiosaurian diversity, and distributions extended to above the Arctic Circle. In the Southern Hemisphere, high-latitude occurrences of Cretaceous plesiosaurs have been reported from Seymour Island of the Antarctica Peninsula, New Zealand, Argentina, and Australia. In addition to the new Axel Heiberg occurrence, high-latitude occurrences of plesiosaurs in the Cretaceous of the Northern Hemisphere include reports of plesiosaurs from the Kanguk Formation of Ellesmere Island and from Upper Cretaceous sediments on Banks Island, Eglinton Island and Melville Island in the western Canadian Arctic. The ages of some of the western Arctic occurrences are not yet well constrained, but it is possible that they are from sedimentary correlates of the Kanguk Formation. Seasonal migration of vertebrates has been discussed for some Cretaceous Arctic vertebrates. But the high-latitude distribution of plesiosaurs may also be a further reflection of the extreme Turonian-Coniacian climatic warmth, which is apparent from other vertebrates found in the Axel Heiberg assemblage” – Deborah Vandermark et al., Late Cretaceous Plesiosaur Teeth from Axel Heiberg Island, Nunavut, Canada,Arctic 59 (1) (2006), pp.79-82
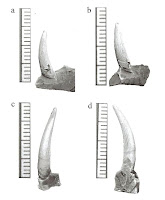

(Left) Freshwater Elasmosaurid Teeth, Axel Heilberg Island, High Canadian Arctic (from Vandermark et al. 2006) (Right) Leatherback Turtle (Dermochelys coracea)
“There used to be a time when the BBC’s ability to produce good science documentaries was highly respected: they generally portrayed a balanced view of the subject, had their facts straight, and were careful not to cast ideas, or scientists themselves, in an inappropriate light. Alas, those days are long gone, as viewing of any Horizon documentary made within the last five years will show. The BBC’s Loch Ness documentary shown last Sunday was no exception, though let me say that I like Steve Leonard and I enjoy watching his stuff. When wondering if a hypothetical plesiosaur could survive in waters as cold as those in Loch Ness, Leonard pointed to leatherbacks and to Australian fossil sites yielding (apparently) coldwater plesiosaurs.
Leatherbacks really can swim in cold boreal seas , but despite initial results from Paladino et al. (1990) they are no longer thought to have a resting metabolic rate elevated compared to that of other similar-sized reptiles. In other words, they are bradymetabolic. However, they are endotherms and have a bunch of features that allow them to be cold tolerant including an insulative carapace, a thick, oil-saturated skin, fibrous fatty tissue and counter current heat exchange mechanisms in the flippers. The presence or absence of all these things can be determined from fossils and there are enough good plesiosaur fossils (including at least one good specimen with skin impressions- my info on it is from Arthur Cruikshank, an expert on plesiosaur anatomy) for us to be confident that these things were absent in plesiosaurs. The bone microtexture and histology of plesiosaurs is not consistent with endothermy- they appear to have been ‘traditional’ reptiles. So the only getout for having them as cold tolerant is to argue that they were gigantotherms.
Problem: the entire theory of gigantothermy is based on Paladino et al.’s erroneous data on leatherbacks, and it has since been retracted by these authors. Given that other studies do not show leatherbacks to have an elevated metabolic rate (Lutcavage et al. 1990), the theory of gigantothermy has died a death, and there is no evidence that giant bradymetabolic vertebrates converge in physiology with giant tachymetabolic ones. Even if gigantothermy were a viable theory, in marine reptiles it is only theoretically possible with the suite of features cited above. Indeed Orenstein (2001) writes ‘Gigantothermy…would not be enough to keep a leatherback warm in cold northern waters’ (pg. 134). At the moment the idea that plesiosaurs may have been cold tolerant is not based on any good evidence.
As for the evidence showing that some Cretaceous plesiosaurs inhabited cold water with icebergs etc., this is controversial: the Cretaceous poles were nowhere near as cold as the modern ones and the evidence that the Australian sites they referred to were frequented by icebergs rests on the presence of drop stones- rocks alien to the local sedimentary geology and which appear to have been carried to their new home by ice. The problem is that icebergs are not the only way in which drop stones get dropped. Stones and rocks can also be carried for miles and miles in the roots of floating trees and as seaweed holdfasts.”- Darren Naish, CZ Conversations: Darren Naish on Plesiosaurs, Basilosaurs and the Problems with Reconstructions, North American BioFortean Review 5 (3) (2003), pp.10-19
“ Higher metabolic rates for these ancient reptiles, relative to modern ones, have previously been suggested, on the basis of bone histology and swimming energetics.All three groups (plesiosaurs, ichthyosaurs and mosasaurs) had a higher body temperature than co-occurring fish by about 5° to 20°C, with the exception of Triassic ichthyosaurs. This suggests that they had heat conservation systems such as blubber layers and specialized blood circulation.”- Ryosuke Motani, Warm-Blooded “Sea Dragons”?, Science 328 (2010), pp.1361-1362
“ No other known extant reptile shows this combination of chondro-osseus developmental features. However, certain extinct reptiles show some similarities to the leatherback: plesiosaurs have been described as having endochondral and periosteal cones that do not remodel ; ichthyosaurs, plesiosaurs, nothosaurs and mesosaurs are known to have amedullary bones with cancellous-compacta differentiation paralleling that of marine mammals, and protostegid turtles have also been noted briefly to be somewhat similar in this respect. All these fossil reptiles were highly adapted to a marine existence as shown by other skeletal features. No terrestrial vertebrate has these chondro-osseus developmental features. The fact that such diverse groups as cetaceans, sirenians, pinnipeds, penguins, extinct marine reptiles and amphibians, and leatherback turtles have such a high degree of physical similarity in bone morphology suggests an underlying mechanism of marine adaptability which has led to a highly developed pattern of skeletal evolutionary convergence .”-Anders G. J. Rhodin et al., Chondro-osseus Morphology of Dermochelys coreacea, a marine reptile with mammalian skeletal features, Nature 290 (5803) (1981), pp. 244-246
 |
The Plesiosaur Polycotylus latippinus giving birth
|
“Among modern reptiles, the plesiosaur-like trait combination of viviparity, small brood size, and large birth size is rare, but it does occur in the scincid Egernia species group. Because both cetaceans and Egernia-group lizards are highly social and engage in substantial maternal care, plesiosaurs may have behaved similarly. We hypothesize that large plesiosaur fetus size may indicate that plesiosaurs lived in gregarious social groups and engaged in parental
care. ”- F. R. O’Keefe and L. M. Chiappe, Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia),Science 333(2011), pp. 870-873
“ The ability of plesiosaurs to move on land is another point of contention . The mechanics of their skeletons imply a completely aquatic existence: the limb girdles are only weakly connected to the axial skeleton and this would inhibit the transfer of force from limb strokes into movement on land. However, small plesiosaurs may have been relatively unaffected by these constraints and might have used their powerful limb downstrokes to propel themselves forward in short ‘hops’.”- Adam Stuart Smith, Fossils Explained 54: Plesiosaurs, Geology Today 24(2) (2008), pp.71-75
The plesiosaur Cryptocleidus oxoniensis on land from the BBC’s Walking With Dinosaurs, Copyright Dave Martill and Darren Naish 2000
Arthur Grant’s sketch of Loch Ness “ Monster” on land, Jan. 1934
“An hypothesis requires two elements. It has to be 1) verifiable - supported by evidence and 2) falsifiable - capable of being shown false by evidence. For example: "Plesiosaurs became extinct at the end of the Cretaceous period" is a useful hypothesis. It is verified by the fact that we have not found any plesiosaurs in post-Cretaceous deposits. It can be falsified by the discovery of plesiosaurs in post-cretaceous deposits. So far, no plesiosaur fossils have been found in post-cretaceous deposits, so the hypothesis stands. "Plesiosaurs did not become extinct at the end of the Cretaceous period" sounds as if it is the corollary hypothesis to the first hypothesis. After all, if they didn't become extinct at the end of the cretaceous, the only possible alternative is that they did not become extinct. But science can't work this way: The assertion is not verified - there are no plesiosaur fossils in post-cretaceous deposits, even though it could be verified by the discovery of such fossils. However, it cannot be falsified: there is no evidence which could show that plesiosaurs did *not* become extinct at the end of the cretaceous. We may look for such fossils but never find them because they don't exist, or because we are looking in the wrong places, or because the places in which post-cretaceous plesiosaurs lived did not provide conditions suitable for fossilisation, or for any other number of unknown reasons. This makes is it not a useless hypothesis, but not an hypothesis at all. An inherent element of any hypothesis that it should be falsifiable. Without falsifiability there is no hypothesis.
There is an obvious parallel in the discovery of Latimeria. Before 1938, one might have formulated the hypothesis that coelacanths became extinct by the end of the Cretaceous. This hypothesis would be verified by the lack of such fossils, and could be falsified by the discovery of post-cretaceous coelacanths. It was falsified rather dramatically in 1938 when Latimeria was discovered. This does not mean that it was not a good hypothesis - the fact that it was falsified means that it was. Furthermore, it does not mean that we now have to work from the hypothesis that coelacanths survived to the present day: that's a fact, not an hypothesis. The assertion that they have survived to the present day is verified by the existence of Latimeria, but that assertion is not an hypothesis: it cannot be falsified. Latimeria can't be ‘undiscovered’ again.”- Richard Forrest, Something about hypotheses,

















































{kind=link}